2017 mikrobiaalisista lektiineistä
Microbial lectins http://nizetlab.ucsd.edu/publications/Lectins-Chapter37.pdf Microbial Lectins: Hemagglutinins,Adhesins, and Toxins Viruses, bacteria, fungi, and protozoa express an enormous array of glycan-binding proteins, also called lectins. Many of these microbial lectins were originally detected based on their ability to aggregate or induce the hemagglutination of red blood cells (erythrocytes). The first microbial hemagglutinin identified was isolated from the influenza virus, and it was shown by Alfred Gottschalk in the early 1950s to bind erythrocytes and other cells via the sialic acid component of host cell-surface glycoconjugates. Don Wiley and associates crystallized the influenza hemagglutinin and determined its structure in 1981. Later they solved the structure of hemagglutinin cocrystals bound to sialyllactose, providing molecular insight into the affinity and specificity of the receptor ligand binding sites. Since then, a number of viral hemagglutinins have been identified and structurally elucidated. Nathan Sharon and colleagues first described bacterial surface lectins in the 1970s. Their primary function is to facilitate the attachment or adherence of bacteria to host cells, a prerequisite for bacterial colonization and infection (Chapter 42). Thus, bacterial lectins are often called adhesins , and these bind corresbondingglycan receptorson the surface of the host cells viacarbohydrate-recognition domains (CRDs) (“receptor” in this case is equivalent to “ligand” for animal cell lectins). Like animal lectins, some microbial adhesins bind to terminal sugar residues via the CRD, whereas others bind to internal sequences found in linear or branched oligosaccharide chains. The interaction of adhesins with host glycans is an important determinant of the tropism of the corresponding pathogen or symbiont. Detailed studies of the specificity of such microbial lectins have led to the identification and synthesis of powerful inhibitors of adhesion that may form the basis for novel therapeutic agents to combat infectious disease
(Chapter 42
2017 N-glykaanivälitteinen adheesio vahvistuu patogeenin ja reseptorin välisessä sitoutumisessa https://www.nature.com/articles/s41598-017-07220-w N-glycan mediated adhesion strengthening during pathogen-receptor binding revealed by cell-cell force spectroscopy
Inflammasomes
are multimeric protein complexes that assemble in the cytosol of many
types of cells, including innate immune cells. The inflammasomes can be
activated in response to infection or in response to stress signals that
induce damage in the host cell membranes.
These platforms trigger
inflammatory processes, cell death, and the control of microbial
replication. Many inflammasomes have been described so far, including
NLRP3,
NAIP/NLRC4,
caspase-11, and
AIM2 (absent in n melanoma 2)
Preferred Names
interferon-inducible protein AIM2
The ligand forNLRP3 is still unidentified, but the efflux of K+ is essential for NLRP3
activation.
By contrast, inflammasomes, such as those composed of
NAIP/NLRC4, caspase-11, and AIM2, can be activated by bacterial
flagellin, LPS, and dsDNA.
The knowledge of inflammasome biology has
advanced tremendously in the last decade, fostered by the use of model
organisms, such as Legionella pneumophila
This bacterium evolved, infecting unicellular protozoa in freshwater
environments, and the human infection is accidental. Thus, L. pneumophila
did not evolve sophisticated mechanisms to inhibit mammalian innate
immunity.
For this reason, it has emerged as a very appropriate model of
a pathogenic microbe for the investigation of inflammasome biology.
In
this review, we highlight the current information regarding the biology
of inflammasomes and emphasize the advances achieved using L. pneumophila
We also describe the inflammasomes activated in response to L. pneumophila infection and discuss the effector mechanisms that operate to clear the infection.
Etsin ensin NOD -kaltaisten reseptorien ja Legionellan yhteisiä artikkeleita ja niistä otan tähän sitaatin.
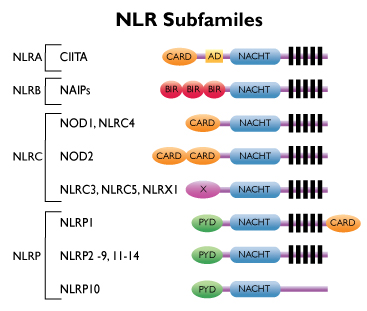
(Mitä NOD reseptorit ja niiden kaltaiset reseptorit ovat? Niistä on arikkeli mm tässä lähteessä: https://www.invivogen.com/review-nlr
Toll-reseptorit solupinnalla tunnetaan tarkemmin niitä on ainakin 9. Ne ovat patogeenin tai vaaran hahmontunnistaja reseptoreita. Näitä NOD reseptoreita on tulehtuneen solun sisällä. NLR: 22 kpl). Koska Legionella on solunsisällä replikoituva, etsin sen suhteita NOD- reseptoreihin. Löysin 21 viitettä.
Park B, Park G, Kim J, Lim SA, Lee KM. Arch Pharm Res. 2017 Feb;40(2):131-145. doi: 10.1007/s12272-016-0859-9. Epub 2017 Jan 6. Review. PMID:28063015
Tiivistelmästä suomennosta.
Legionella pneumophila on eräs etiologinen agenssi vaikeassa legionelloosi-keuhkotulehduksessa. Tämä gram-negatiivinen bakteeri replikoi luonnossa erilaisissa makeanveden ameeboissa, mutta myös ihmisen keuhkoalveolien makrofageissa. Isäntäsolun sisällä Legionella indusoi ei-endosomaalisten replikaatiokykyisten fagosomien tuotannon injisoimalla effektoriproteiinejaan solun sytosoliin .
Luonnolliset immuunivasteet ovat eturintamapuolustusta (=valmiina olevaa resurssia) Legionellaa vastaan infektion varhaisvaiheessa ja tekee eroa Legionellan ja isäntäsolun välillä käyttämällä taudinaiheuttajan hahmon tunnistavia (PAMP) reseptoreita (PRR) kuten TOLL-reseptorin (TLR) tai NOD- reseptorin kaltaisia (NLR) tai tai RIG-1 kaltaisia reseptoreita ( RLR). Ne pystyvät havaitsemaan patogeeneihin liittyviä molekulaarisia hahmoja (PAMP), joita kehossa , isäntäsolussa itsessään ei ole.
Keuhkojen Legionella- tulehduksen aikana rekrytoituu keuhkoihin useita erilaisia tulehdussoluja kuten makrofageja, neutrofiilejä, luonnollisia tappajasoluja (NK) , suuria mononukleaarisia soluja , B-imusoluja ja T-imusoluja: sekä CD4+ että CD8+ T-lymfosyyttisoluja ja ensisijaisesti ne asettuvat kudossolujen väleihin, interstitiaalisesti, kontrolloimaan Legionellaa.
Keuhkoissa menossa olevissa Legionella-infektioissa myös eri sytokiinien ja kemokiinien väliset vuorovaikutukset moduloivat isäntäkehon immuunivastetta . NK-soluilla tapahtuvasta tunnistuksesta liipaistuu esiin kehon omia effektorifunktioita kuten sytokiinien ja kemokiinien erittymistä ja niistä aiheutuu kohteena olevien solujen lyysi, hajoaminen. NK- solujen, dendriittisolujne, monosyyttien ja makrofagien keskeinen vuorovaikutus antaa pääasiallisen etulinjapuolustuksen Legionellaa vastaan, kun taas spesifinen immuunivaste, T- ja B-solujen aktivoituminen johtaa lopulta infektion päättymiseen ja pystyttää Legionella-spesifistä muistijälkeä isäntäkehoon.
Abstract
Legionella
pneumophila is an etiological agent of the severe pneumonia known as
Legionnaires' disease (LD). This gram-negative bacterium is thought to
replicate naturally in various freshwater amoebae, but also replicates
in human alveolar macrophages. Inside host cells, legionella
induce the production of non-endosomal replicative phagosomes by
injecting effector proteins into the cytosol. Innate immune responses
are first line defenses against legionella during early phases of infection, and distinguish between legionella and host cells using germline-encoded pattern recognition receptors such as Toll-like receptors , NOD-like receptors, and RIG-I-like receptors, which sense pathogen-associated molecular patterns (PAMP) that are absent in host cells.
During pulmonary legionella infections, the interplay between distinct cytokines and chemokines also modulates innate host responses to clear legionella
from the lungs. Recognition by NK cell receptors triggers effector
functions including secretion of cytokines and chemokines, and leads to
lysis of target cells. Crosstalk between NK cells and dendritic cells,
monocytes, and macrophages provides a major first-line defense against legionella infection, whereas activation of T and B cells resolves the infection and mounts legionella-specific memory in the host.
4. Yksi L. pneumophilan effektoreista on LegS2, SP1-lyaasi
(Kommentti: Tällainen entsyymikuuluu ihmisenkin Sfingomyeliiniaineenvaihdunnan terminaaliseen kohtaan membraani remodelling kierrossa ) . Lpn-mutantti, jolta tämä funktio puuttuu ilmentää isäntäkehossa atyyppisia mitokondrioita. Bakterin effektori ilmeisesti pehmentää mitokondrian ulkokalvonrakennetta ja saa mitokondriat jopa fusoitumaan ja näyttämään amorfisilta).
The Sphingosine-1-Phosphate Lyase (LegS2) Contributes to the Restriction of Legionella pneumophila in Murine Macrophages. Abu Khweek A, Kanneganti A, Guttridge D DC, Amer AO.PLoS One. 2016 Jan 7;11(1):e0146410. doi: 10.1371/journal.pone.0146410. eCollection 2016. PMID: 26741365 L. pneumophila is the causative agent of
Legionnaires’ disease, a human illness characterized by severe
pneumonia. In contrast to those derived from humans, macrophages derived
from most mouse strains restrict L. pneumophila replication. The restriction of L. pneumophila
replication has been shown to require bacterial flagellin, a component
of the type IV secretion system as well as the cytosolic NOD-like
receptor (NLR) Nlrc4/ Ipaf. These events lead to caspase-1 activation
which, in turn, activates caspase-7. Following caspase-7 activation, the
phagosome-containing L. pneumophila fuses with the lysosome, resulting in the restriction of L. pneumophila
growth. The LegS2 effector is injected by the type IV secretion system (T4SS)
and functions as a sphingosine 1-phosphate lyase. It is homologous to
the eukaryotic sphingosine lyase (SPL), an enzyme required in the
terminal steps of sphingolipid metabolism. Herein, we show that mice
Bone Marrow-Derived Macrophages (BMDMs) and human Monocyte-Derived
Macrophages (hMDMs) are more permissive to L. pneumophila legS2 mutants than wild-type (WT) strains. This permissiveness to L. pneumophila legS2
is neither attributed to abolished caspase-1, caspase-7 or caspase-3
activation, nor due to the impairment of phagosome-lysosome fusion.
Instead, an infection with the legS2 mutant resulted in the
reduction of some inflammatory cytokines and their corresponding mRNA;
this effect is mediated by the inhibition of the nuclear transcription
factor kappa-B (NF-κB). Moreover, BMDMs infected with L. pneumophila legS2
mutant showed elongated mitochondria that resembles mitochondrial
fusion. Therefore, the absence of LegS2 effector is associated with
reduced NF-κB activation and atypical morphology of mitochondria. Free PMC Article Similar articles
5. Inflammasomi on multiproteiinikompleksi, johon kuuluu jäseninä NOD:n kaltianen reeptoriperhe(NLR) ja kaspaasi-1.
Caspase-11
and caspase-1 differentially modulate actin polymerization via RhoA and
Slingshot proteins to promote bacterial clearance. Caution K, Gavrilin MA, Tazi M, Kanneganti A, Layman D, Hoque S, Krause K, Amer AO. Sci Rep. 2015 Dec 21;5:18479. doi: 10.1038/srep18479. Inflammasomes are multiprotein complexes that include members of the NOD-likereceptor family and caspase-1. Caspase-1 is required for the fusion of the Legionella
vacuole with lysosomes. Caspase-11, independently of the inflammasome,
also promotes phagolysosomal fusion. However, it is unclear how these
proteases alter intracellular trafficking. Here, we show that caspase-11
and caspase-1 function in opposing manners to phosphorylate and
dephosphorylate cofilin, respectively upon infection with Legionella.
Caspase-11 targets cofilin via the RhoA GTPase, whereas caspase-1
engages the Slingshot phosphatase. The absence of either caspase-11 or
caspase-1 maintains actin in the polymerized or depolymerized form,
respectively and averts the fusion of pathogen-containing vacuoles with
lysosomes. Therefore, caspase-11 and caspase-1 converge on the actin
machinery with opposing effects to promote vesicular trafficking.
PMID:26686473 Free PMC Article
In
contrast to TLR5, NLRC4 did not regulate recruitment of neutrophils to
the lung. Lp replication in murine alveolar macrophages was NLRC4-dependent
and TLR5-independent. These studies reveal
that NLRC4 and TLR5 mediate different roles in the inflammatory response
to Lp flagellin in an aerosolized infection model and NLRC4 regulates
replication in both lungs and alveolar macrophages.Free PMC Article Similar articles
7. NOD-reseptorit inflammasomissa, joka on laaja kompleksi sytoplasmassa, tunnistaa mikrobitulehduksen ja vaaran molekyylejä ja indusoi kaspaasi-1-aktivaatiosta riippuvan sytokiinituotannon ja makrofagin tulehduksellisen kuoleman ( bakteerimassan kanssa lyysissä). NLRC4 tunnistaa bakteeriflagelliinia ja T4SS sekreetiosysteemin.
Nature. 2011 Sep 14;477(7366):596-600. doi: 10.1038/nature10510. PMID:21918512 Inflammasomes are large cytoplasmic complexes that sense microbial
infections/danger molecules and induce caspase-1 activation-dependent
cytokine production and macrophage inflammatory death. The inflammasome
assembled by the NOD-likereceptor
(NLR) protein NLRC4 responds to bacterial flagellin and a conserved
type III secretion system (TTSS) rod component. How the NLRC4
inflammasome detects the two bacterial products and the molecular
mechanism of NLRC4 inflammasome activation are not understood. Here we
show that NAIP5, a BIR-domain NLR protein required for Legionella
pneumophila replication in mouse macrophages, is a universal component
of the flagellin-NLRC4 pathway. NAIP5 directly and specifically
interacted with flagellin, which determined the inflammasome-stimulation
activities of different bacterial flagellins. NAIP5 engagement by
flagellin promoted a physical NAIP5-NLRC4 association, rendering full
reconstitution of a flagellin-responsive NLRC4 inflammasome in
non-macrophage cells. The related NAIP2 functioned analogously to NAIP5,
serving as a specific inflammasome receptor
for TTSS rod proteins such as Salmonella PrgJ and Burkholderia BsaK.
Genetic analysis of Chromobacterium violaceum infection revealed that
the TTSS needle protein CprI can stimulate NLRC4 inflammasome activation
in human macrophages. Similarly, CprI is specifically recognized by
human NAIP, the sole NAIP family member in human. The finding that NAIP
proteins are inflammasome receptors for bacterial flagellin and TTSS
apparatus components further predicts that the remaining NAIP family
members may recognize other unidentified microbial products to activate
NLRC4 inflammasome-mediated innate immunity.Similar articles
8. NOD-perheen reseptorista NAIP5 inflammasomissa. Rajoittaa Legionellaa ja tunnistaa flagelliinia.
Global cellular changes induced by Legionella pneumophila infection of bone marrow-derived macrophages. Fortier A, Faucher SP, Diallo K, Gros P. Immunobiology. 2011 Dec;216(12):1274-85. doi: 10.1016/j.imbio.2011.06.008. Epub 2011 Jun 30. PMID:21794945
The nucleotide-binding oligomerization domain (Nod)-likereceptor(NLR) family member Naip5 plays an essential role in restricting Legionella pneumophila growth inside primary macrophages. Upon interaction with bacterial flagellin, the intracellular receptorNaip5 forms a multi-proteincomplex, the inflammasome, which activation
has a protective role against infection.
9. Legionella pneumophila manipuloi tehottomiksi kaksi isäntäkehon apoptoottista tietä: sekä kanonisen apoptoottisen mitokondriaalisen tien että pyroptoottisen NOD:in kaltaisten reseptorien kontrolloiman inflammasomisen tien.
Programmed cell
death is considered the ultimate solution for the host to eliminate
infected cells, leading to the abolishment of the niche for microbial
replication and the ablation of infection. Thus, it is not surprising
that successful pathogens have evolved diverse strategies to reprogram
the cell death pathways for their proliferation. Using effector proteins
translocated by the Dot/Icm type IV secretion system, the facultative
intracellular pathogen Legionella
pneumophila manipulates multiple host cellular processes to create a
niche within host cells to support its replication. Investigation in the
past decade has established that in mammalian cells this bacterium
actively modulates two host cell death pathways, namely the canonical
apoptotic pathwaycontrolled by the mitochondrion and the pyroptotic
pathway controlled by the Nod-likereceptor
Naip5 and the Ipaf inflammasome. In this review, I will discuss the
recent progress in understanding the mechanisms the bacterium employs to
interfere with these host cell death pathways and how such modulation
contribute to the intracellular life cycle of the pathogen.Similar articles
10. Lpn- infektiossa ASC ja NLRC4 alassäätyneinä monosyyteissä.
J Biol Chem. 2011 Feb 4;286(5):3203-8. doi: 10.1074/jbc.M110.197681. Epub 2010 Nov 19. PMID:21097506 The ability of Legionella
pneumophila to cause pneumonia is determined by its capability to evade
the immune system and grow within human monocytes and their derived
macrophages. Human monocytes efficiently activate caspase-1 in response
to Salmonella but not to L. pneumophila. The molecular mechanism for the
lack of inflammasome activation during L. pneumophila infection is
unknown. Evaluation of the expression of several inflammasome components
in human monocytes during L. pneumophila infection revealed that the
expression of the apoptosis-associated speck-like protein (ASC) and the NOD-likereceptor
NLRC4 are significantly down-regulated in human monocytes. Exogenous
expression of ASC maintained the protein level constant during L.
pneumophila infection and conveyed caspase-1 activation and restricted
the growth of the pathogen. Further depletion of ASC with siRNA was
accompanied with improved NF-κB activation and enhanced L. pneumophila
growth. Therefore, our data demonstrate that L. pneumo phila manipulates
ASC levels to evade inflammasome activation and grow in human monocytes.
By targeting ASC, L. pneumophila modulates the inflammasome, the
apoptosome, and NF-κB pathway simultaneously.Free PMC Article Similar articles
11. PAMP reseptorit Nod1 ja Nod2 vastaavat neutrofiilien rekrytoimisesta hiiren keuhkoon legionella pneumophila-infektiossa.
Legionella
pneumophila is the causative agent of Legionnaires' disease, a serious
and often fatal form of pneumonia. The susceptibility to L. pneumophila
arises from the ability of this intracellular pathogen to multiply in
human alveolar macrophages and monocytes. L. pneumophila also replicates
in several professional and non-professional phagocytic human-derived
cell lines. With the exception of the A/J mouse strain, most mice
strains are restrictive, thus they do not support L. pneumophila
replication. Mice lacking the NOD-likereceptor
Nlrc4 or caspase-1 are also susceptible to L. pneumophila. On the other
hand, in the susceptible human hosts, L. pneumophila utilizes several
strategies to ensure intracellular replication and protect itself
against the host immune system. Most of these strategies converge to
prevent the fusion of the L. pneumophila phagosomewith the lysosome,
inhibiting host cell apoptosis, activating survival pathways, and
sequestering essential nutrients for replication and pathogenesis. In
this review, we summarize survival mechanisms employed by L. pneumophila
to maintain its replication in human cells. In addition, we highlight
different human-derived cell lines that support the multiplication of
this intracellular bacterium. Therefore, these in vitro models can be
applicable and are reproducible when investigating L.
pneumophila/phagocyte interactions at the molecular and cellular levels
in the human host.KEYWORDS: NOD-like receptors; Toll-like receptors; neuronal apoptosis-inhibitory proteins; pathogen-associated molecular patterns Free PMC Article Similar articles
13 Legionella pn. moduloi kaspaaseja ja saa ne toimimaan ei-apoptoottisina.
Legionella pneumofilasta on tullut mallijärjestelmä kaspaasien ei-apoptoottisten funktioiden ja immuunitehtävien ratkaisemiseen. Sallivissa soluissa Lpn-vakuolit (LCV) välttävät endosomaalisia kuljetusteitä ja endoplasminen verkosto(ER) muokaa vakuoleja uuteen muotoon ( jossa replikaatio voi tapahtua). Endosomitien evaasio välittyy Legionellan Dot/Icm- tyyppi4-sekreetiosysteemillä (T4SS). Lpn- infektiossa flagelliinia tunnistaa NOD:n kaltainen reseptori NLRC4 (IPAF) , mikä johtaa kaspaasi-1 aktivaatioon inflammasomikompleksissa. NLRC4 inflammasomin alavirrassa aktivoituu kaspaasi-7 ja edistää ei-apoptoottisia funktioita, kuten fagosomin kypsymistä ja bakteriaalista hajoittamista . On tehty mielenkiintoinen havainto kaspaasi-3:n aktivoitumisesta infektion varhaisvaiheissa permissiivisissä soluissa, sillä se ei johdakaan apoptoosiin ja niin on aivan infektoitumisen myöhäisvaiheisiin asti; tähän on syynä Dot/Icm:n välittämät antiapoptoottiset signaalit, jotka tekevät infektoituneet solut resistenteiksi ulkopuolisille apoptoosin aiheuttajille. Sen takia kaspaasi 1:n ja exekutiivisten kaspaasien ei-apoptoottiset funktiot ovat temporaalisesti ja spatiaalisesti moduloituja Lpn - infektion aikana, mistä määräytyy permissiivisyys (sallivuus) solunsisäiseen bakteerireplikoitumiseen. Artikkelissa tehdään selkoa L. pneumophilan uusista kaspaasi- aktivoitumisteistä ja pohditaan niiden osuutta geneettisessä restriktiossa ja infektio- permissiivisyydessä.
Amer AO. Cell Microbiol. 2010 Feb;12(2):140-7. doi: 10.1111/j.1462-5822.2009.01401.x. Epub 2009 Oct 27. Review. PMID: 19863553Legionella
pneumophila has become a model system to decipher the non-apoptotic
functions of caspases and their role in immunity. In permissive cells,
the L. pneumophila-containing vacuole evades endosomal traffic and is
remodelled by the endoplasmic reticulum. Evasion of the endosomes is
mediated by the Dot/Icm type IV secretion system. Upon L. pneumophila
infection of genetically restrictive cells such as wild-type (WT)
C57Bl/6J murine macrophages, flagellin is sensed by the NOD-likereceptor
Nlrc4 leading to caspase-1 activation by the inflammasome complex.
Then, caspase-7 is activated downstream of the Nlrc4 inflammasome,
promoting non-apoptotic functions such as L. pneumophila-containing
phagosome maturation and bacterial degradation. Interestingly, caspase-3
is activated in permissive cells during early stages of infection.
However, caspase-3 activation does not lead to apoptosis until late
stages of infection because it is associated with potent
Dot/Icm-mediated anti-apoptotic stimuli that render the infected cells
resistant to external apoptotic inducers. Therefore, the role of
caspase-1 and non-apoptotic functions of executioner caspases are
temporally and spatially modulated during infection by L. pneumophila,
which determine permissiveness to intracellular bacterial proliferation.
This review will examine the novel activation pathways of caspases by
L. pneumophila and discuss their role in genetic restriction and
permissiveness to infection.Similar articles
14.Mitä tarvitaan legionellan restriktioon makrofagissa.
16. NLR-perheenjäsen NLRC4 eli IPAF omaa CARD-domeenin ja NACHT- domeenin. Intrasellulaari Gram- negat. bakteeri indusoi 1-kaspaasin IPAF välitteisesti ja infektoituneen makrofagin kuoleman ja proinflammatorisia sytokiineja. ( Useimmat ihmiset eivät sairastu Legionella-bakteeriin, kun immuunivaste toimii normaalisti, tai paremminkin: useimmat ihmiset toipuvat Legionellasta, joka on diagnosoitu).
The NOD-likereceptor
(NLR) family of proteins is involved in the regulation of innate immune
responses and cell death pathways. Recent findings show that the NLR
family member NLRC4 (also known as IPAF) has important roles in innate
immune responses to Gram-negative bacteria. Macrophages infected with Legionella
pneumophila, Salmonella typhimurium, Shigella flexneri, or Pseudomonas
aeruginosa activate caspase-1 in an NLRC4-dependent manner leading to
macrophage cell death and the release of proinflammatory cytokines. This
review will discuss these findings as well as the role of bacterial
type III and type IV secretion systems and flagellin in NLRC4-mediated
caspase-1 activation.Free PMC Article Similar articles
Semin Immunopathol. 2007 Sep;29(3):275-88. Epub 2007 Aug 10. Review.PMIDAbstractThe innate immune
system precisely modulates the intensity of immune activation in
response to infection. Flagellinis a microbe-associated molecular
pattern that is present on both pathogenic and nonpathogenic bacteria.
Macrophages and dendritic cells (DC) are able to determine the virulence of
flagellated bacteria by sensing whether flagellin remains outside the
mammalian cell, or if it gains access to the cytosol. Extracellular
flagellin is detected by TLR5, which induces expression of
proinflammatory cytokines, while flagellin within the cytosol of
macrophages is detected through the Nod-likereceptor (NLR) Ipaf, which activates caspase-1. In macrophages infected with Salmonella typhimurium or Legionella
pneumophila, Ipaf becomes activated in response to flagellin that
appears to be delivered to the cytosol via specific virulence factor
transport systems (the SPI1 type III secretion system (T3SS) and the
Dot/Icm type IV secretion system (T4SS), respectively). Thus, TLR5
responds more generally to flagellated bacteria, while Ipaf responds to
bacteria that express both flagellin and virulence factors
21. Intrasellulaarisen Legionella pneumophila-kasvun restriktioon vaaditaan Ipaf:ista riippuva kaspaasi-1 aktivaatio ja funktionaalinen NAIP5-signalointi.
The Nod-likereceptor family member Naip5/Birc1e restricts Legionella pneumophila growth independently of caspase-1 activation.Lamkanfi M, Amer A, Kanneganti TD, Muñoz-Planillo R, Chen G, Vandenabeele P, Fortier A, Gros P, Núñez G.J Immunol. 2007 Jun 15;178(12):8022-7. PMID: 17548639Abstract Similar to Ipaf and caspase-1, the Nod-likereceptor protein Naip5 restricts intracellular proliferation of Legionella
pneumophila, the causative agent of a severe form of pneumonia known as
Legionnaires' disease. Thus, Naip5 has been suggested to regulate Legionella
replication inside macrophages through theactivation of caspase-1. In
this study, we show that cytosolic delivery of recombinant flagellin
activated caspase-1 in A/J macrophages carrying a mutant Naip5 allele,
and in C57BL/6 (B6) macrophages congenic for the mutant Naip5 allele
(B6-Naip5(A/J)), but not in Ipaf(-/-) cells. In line with these results,
A/J and B6-Naip5(A/J) macrophages induced high levels of caspase-1
activation and IL-1beta secretion when infected with Legionella. In addition, transgenic expression of a functional Naip5 allele in A/J macrophages did not alter Legionella-induced
caspase-1 activation and IL-1beta secretion. Notably, defective Naip5
signaling renders B6-Naip5(A/J) macrophages permissive for Legionella proliferation despite normal caspase-1 activation. These results indicate that the restriction of intracellular Legionella
replication is more complex than previously appreciated and requires
both Ipaf-dependent caspase-1 activation as well as functional Naip5
signaling. Free Article Similar articles
Bakteerin vaippa on ratkaisevassa osassa infektiotautioen patogeneesissä. Tässä katsauksessa on yhteenvetoa nykyisestä (2011) tiedosta legionella pneumophila soluvaipan rakenteesta ja molekulaarisesta koostumuksesta. Käsitellään lipopolysakkaridien (LPS) synteesiä kalvon ja periplasmisten proteiinien biologisia aktiivisuuksia ja niiden ratkaisevia funktioita patogeenin ja sen isännän välisessä vuorovaikutuksessa. Bakteerin kiinnittymisen (adheesio) , invasoitumisen ja solun sisällä elossapysymisen lisäksi kiinnitetään erityishuomiota raudanhankintaan, detoksikaatioon, immuunivasteen avainasemassa oleviin esiinsaajiin ja ulomman rakkulakalvon erilaisiin funktioihin. Kirjallisuuden kriittisestä analysoinnista käy ilmi, että legionellasolupinnan dynamiikka ja fenotyyppinen plastisuus erilaisten metabolisten vaiheiden aikana vaatii tulevina aikoina enemmän huomionkiinnittämistä. Avainsanoja: Legionella pneumophila,bakteerivaippa, fosfolipidit, kalvoproteiinit, LPS, ulkokalvo rakkulat.
Abstract The
bacterial envelope plays a crucial role in the pathogenesis of
infectious diseases. In this review, we summarize the current knowledge
of the structure and molecular composition of the Legionella pneumophila
cell envelope. We describe lipopolysaccharides biosynthesis and the
biological activities of membrane and periplasmic proteins and discuss
their decisive functions during the pathogen–host interaction. In
addition to adherence, invasion, and intracellular survival of L. pneumophila,
special emphasis is laid on iron acquisition, detoxification, key
elicitors of the immune response and the diverse functions of outer
membrane vesicles. The critical analysis of the literature reveals that
the dynamics and phenotypic plasticity of the Legionella cell surface during the different metabolic stages require more attention in the future. Keywords: Legionella pneumophila, bacterial envelope, phospholipids, membrane proteins, LPS, outer membrane vesicles.
Bakteerisolukalvolla on useita perusfunktioita. Ne suojaavat bakteeria ympäristön aiheuttamilta vaaroilta. Ne sallivat ravintoaineiden selektiivisen sisäänpäsyn ja spesifisen jätetuotteiden uloskuljetuksen sekä eritysjärjestelmän tuotesubstraattien ulospääsyn solusta. Lisäksi ne välittävät suoraa kontaktia muihin organismeihin. Tämä pitää paikkansa varsinkin patogeenisiin bakteereihin, joiden usein hyvin spesifiset interaktiot isäntäorganismin kanssa riippuvat suurelta osalta juuri niiden pintarakenteista. Niinpä Legionella pneumophilan, fakultatiivisen Gram-negatiivisen intrasellulaarisen bakteerin kyky aiheuttaa legionalaistautia on lähinnä sen soluvaipan kmponenttien ja ominaisuuksien varqassa.
Gram-negatiivisten bakteereiden sytoplasmaa (CP) rajoittaa sisäkalvo (IM). Se on kahden fosfolipidikerroksen muodostama ja siinä on integoituneita ja perifeerisiä proteiineja ja lipoproteiineja. Se omaa metabolisia entsyymejä, hengitysketjun komponentteja ja osia koneistosta, joka hankkii rautaa. (Kuva 11)
Bacterial cell envelopes fulfill several
basic functions: They protect the bacterium from environmental hazards,
they allow a selective passage of nutrients into and a specific export
of waste products and secretion system substrates out of the cell.
Additionally, they mediate the direct contact with other organisms. This
holds particularly true for pathogenic bacteria, whose often highly
specific interactions with host organisms depend largely on their
surface structures. Accordingly, the ability of the Gram-negative
facultative intracellular bacterium Legionella pneumophila to cause Legionnaires’ disease hinges predominantly on the components and characteristics of its cell envelope.The
cytoplasm of Gram-negative bacteria is bordered by the inner membrane.
It consists of a bilayer of two phospholipid leaflets with integral and
peripheral proteins and lipoproteins. It harbors metabolic enzymes,
components of the respiratory chain and parts of the iron acquisition
machinery (Figure (Figure11).
Kuvassa on seuraavia symboleja ja lyhennyksiä:
PAL = peptidoglykaaniin liittynyt lipoprteiini
FeoB= raudan kuljetuskohta
PlaB = Fosfolipaasi A/lysofosfolipaasi A.
MOMP= pääasiallinen ulomman kalvon proteiini .
OM = outer membrane, ulompi kalvo; ( ulompi lipidikaksoiskalvo)
IM = inner membrane, sisempi kalvo ( Sisempi lipidikaksoiskalvo)
Mip = makrofagi-infektiivisyyden vahvasitaja
PP = periplasmisia proteiineja
...
Periplasma (PP) sisältää suhteellisen ohuen keroksen peptidoglykaania ja eri proteiineja. Legionellan peptidoglykaani on vahvasti ristikkäin linkkiytynyttä. Periplasmaan sijoittautuu monet detoksikoivat entsyymit, jotka pystyvät tekemään vaarattomaksi ja hajoittamaan miljöön haitalliset aineet. Legionellan eritekoneistot, jotka läpäisevät kaksi kalvoa, ulottuvat periplasmisen tilan läpi.
The periplasm contains a relatively thin layer of peptidoglycan and different proteins. Legionella peptidoglycan is strongly crosslinked (Amano and Williams, 1983).
The periplasm is the location of many detoxifying enzymes which degrade
harmful substances from the environment. Secretion machineries which
cross two membranes also go through the periplasmic space.
Ulkokalvo on asymmetrinen ja sen sisempi lehti on lähinnä fosfolipideistä ja ulompi lehti lähinnä lipopolysakakrideista (LPS). Siihen asettuu proteiineja, joilla on erilaisia virulenssifunktioita kuten adheesio ( kiinnittyminen) ja isäntäsolun sisään ottaminen. Legionellan LPS omaa ainutlatuisen arkkitehtuurin, erityisesti mitä tulee hydrofobiseen O-antigeeniin.
The
outer membrane is asymmetric with an inner leaflet of mostly
phospholipids and an outer leaflet of mostly lipopolysaccharides (LPS).
It harbors proteins with diverse functions in virulence such as adhesion
and uptake into host cells. Legionella LPS has a unique architecture, particularly concerning the hydrophobic O-antigen.
Tietyntyyppiset pinnan lisukkeet kuten karvat( pili) ja siimat ( flagella) , joita baktgeerin motiliteettiin ja patogeenisyyteen vaaditaan, ankkuroituvat sisäkalvoon ja tunkevat esiin solunulkoiseen tilavuuteen.
Certain
types of surface appendages such as pili and flagella, which are
required for bacterial motility and pathogenicity, are anchored in the
inner membrane and protrude into the extracellular space (Liles et al., 1998; Stone and Abu Kwaik, 1998; Heuner and Steinert, 2003).
Ulkokalvon komponenttien virulenssiominaisuudet ovat erityisen tärkeitä ulkokalvoperäisissä rakkuloiissa (OMV). Kuten useimmat bakteerit niin Legionellakin lehteilee näitä OMV-rakkuloita ulkopinnaltaan (OM). Ulkopintarakkulat ovat pallomaisia ja omaavat (yhden) lipidikaksoiskerroksen ja ulkokerrokselle (OM) ominaisia komponetteja ja periplasmisia proteiineja.
Virulence
properties of outer membrane components are particularly important in
regard to outer membrane vesicles (OMVs). Like most bacteria, L. pneumophila
sheds these vesicles from its outer membrane. OMVs are spherical lipid
bilayers and contain outer membrane components and periplasmic proteins.
L. pneumophila-soluvaipan aktuelli rakenne tutkittiin yksityiskohtaisesti elektronimikroskoopilla (Rodgers et Davey, 1982) pian sen jälkeen, kun bakteeri oli keksitty. Eri menetelmin saatiin visualisoitua kummatkin kalvot (OM, IM) sekä peptidoglykaanikerros ja saatiin elävä kuva kaikista komponenteista, joita nykyään analysoidaan lähinnä biokemiallisesti. Nämä tutkijat olivat myös ensimmäisiä osoittamassa L. pneumophilan OMV- rakkuloiden olemassaolon, vaikka niitä nimitettiin kupliksi, "blebs" ja ne selitettiin kondensoituneiksi karvoihin liittyviksi proteiineiksi tai satunnaisiksi ulkokalvoperäisiksi rakenneproteiineiksi.
The actual structure of the L. pneumophila cell envelope was assessed in detail by electron microscopy shortly after the discovery of the bacterium (Rodgers and Davey, 1982).
Both membranes and the peptidoglycan layer were visualized by different
methods, resulting in vivid images of the components that are,
nowadays, analyzed mostly biochemically. The authors are also the first
to show the existence of OMVs of L. pneumophila, even though
they are termed “blebs” and explained as “condensed pili-related
proteins or random structural proteins of the outer membranes.”
L. pneumophila-morfologian laajan tutkimusken suorittivat Faulkner et Garduno( 2002) . He tekivät hypoteesin useiden moforlogisten varianttein olemassaolosta, ja nisitä jokainen vastasi infektiosyklin tiettyä kasvufaasia tai kehitysastetta. Mielenkiintoinen seikka oli, että esitettiin viisi erilaista vaipparakennetta ja ne vaihtelivat paksuuden, kalvokerrosluvun ja yksittäisten komponenttien elektronitiheyden suhteen. Koska joitakin morfologisia variantteja ilmeni vain intrasellulaarisen kasvun aikana, tutkijat olettivat näiden varianttein kehityksen riippuvan isäntäsolun metaboliiteista. Tämä huomio voi selittää sellaisten muotojen puuttuman nestemäisessä väliaineessa tapahtuvan extrasellulaarisen kasvun aikana. Vielä tarvitsee selityksensä näyteitten valmistusprosessin vaikutus näyteissä muodostuviin artefaktoihin.
An
extensive study of L. pneumophila morphology including envelope architecture was performed by Faulkner and Garduño (2002).
They hypothesize the existence of several morphological variants, each
corresponding to a certain growth phase or stage of the infection cycle.
Interestingly, five different envelope structures are presented which
vary in thickness, number of membrane layers, and electron density of
individual components. As some of the morphological variants only
occurred during intracellular growth, the authors propose that the
development of these variants depends on host metabolites. This notion
can explain the absence of these forms during extracellular growth in
liquid media. The impact of processing artifacts arising during the
preparation of the samples, however, remains to be clarified.
(Tämän artikkelin piiriin ei oteta muualla tarkoin selvitettyjä monia sekreetiojärjestelmiä ja ulkokalvon proteiineja, joilla on merkitystä virulenssissa. Tähän kuuluu T1SS ja kaksois arginiinin translokaatio (Tat) eritys (Lammertyn et Anne, 2004), T2SS (Cianciotto,2009), T4SS ja näihin kuuluvat vastaavat translokoituneet effektorit ( (Ninio et Roy, 2007). Lopuksi: erittyneet fosfolipaasit (Pla) yhdistävät Legionella virulenssin isäntä-lipideihin (Banerji et al. 2008) . Vähemmän huomiota on annettu niille muille Legionella soluvaipan komponetteille, jotka eivät kuulu edellämainittuihin komplekseihin. Tämä katsaus keskiittyy nihin kalvokomponentteihin ja miten ne modifioivat Legionellan virulenssin ominaisuuksia.
Many secretion systems and outer membrane proteins with roles in
virulence have been excellently reviewed elsewhere and are not within
the focus of this work. This includes T1SS and twin-arginine
translocation (Tat) secretion (Lammertyn and Anne, 2004), T2SS (Cianciotto, 2009), T4SS as well as their respective translocated effectors (Ninio and Roy, 2007). Finally, secreted phospholipases connect Legionella virulence to host lipids (Banerji et al., 2008). Less attention was paid to other components of the Legionella
cell envelope which are not part of the aforementioned complexes. This
review concentrates on these envelope components and how they mediate Legionella virulence properties.
L.pneumophilan sisäkalvo(IM. Alkaen sisältä ulospäin ensimmäinen kerros sisäkalvoa(IM) on myös sytoplasminen kalvo tqai plasmakalvo nimeltään. Se on lipidikaksoiskerros ja siihen on integroitunut usean systeemin komponetteja, kuten rataa ottava koenisto, hengitysketju ja detoksikaatiojärjestelmä.
The Inner Membrane of L. Pneumophila
Starting
from the inside and proceeding outward, the first layer is the inner
membrane, also termed cytoplasmic or plasma membrane. It is a lipid
bilayer with integrated components of various systems, including the
iron uptake machinery, the respiratory chain, and the detoxification
system (Table (Table11).
Protein
Molecular function
Role in infection/required for
Reference
FeoB
GTP-dependent Fe(II) transporter
Macrophage killing, full virulence in mouse
Petermann et al. (2010), Robey and Cianciotto (2002)
Intracellular
replication in human macrophages, growth under iron-limiting
conditions, cytochrome c-dependent respiration, export of PLC activity
to supernatant
Legionelloosiongelmat pysyvät uutisisia. Instituutioissa ja isoissa rakennuksissa tällaisten vesijärjestelmissä pesivien infektioiden estäminen vaatii omat ohjelmansa
Curr Opin Infect Dis. 2018 Aug;31(4):325-333. doi: 10.1097/QCO.0000000000000468.
Legionellosis remains an important public health threat. To prevent
these infections, staff of municipalities and large buildings must
implement effective water system management programs that reduce Legionella
growth and transmission and all Medicare-certified healthcare
facilities must have water management policies. In addition, we need
better methods for detecting Legionella in water systems and in clinical specimens to improve prevention strategies and clinical diagnosis.
Legionella pneumophila omaa ihmismitokondrian poriinin kaltaisen, jännitteestä riippuvan joniselektiivisen proteiinin, Lpg1974. Se voi muodostaa lipidikaksoiskalvoon jonia selektiivisesti läpäisevän aukon.
.... Genome analyses have shown the presence of genes coding for eukaryotic like proteins in several Legionella
species. The presence of these proteins may assist L. pneumophila in
its adaptation to the eukaryotic host. We studied the characteristics of
a protein (Lpg1974) of L. pneumophila that shows remarkable homologies
in length of the primary sequence and for the identity/homology of many
amino acids to the voltage dependent anion channel (human VDAC1, Porin
31HL)of humanmitochondria. Two different forms of Lpg1974 were
overexpressed in Escherichia coli and purified to homogeneity: the one
containing a putative N-terminal signal sequence and one without it.
Reconstituted protein containing the signal sequence formed
ion-permeable pores in lipid bilayer membranes with a conductance of
approximately 5.4 nS in 1 M KCl. When the predicted N-terminal signal
peptide of Lpg1974 comprising an α-helical structure similar to that at
the N-terminus of hVDAC1 was removed, the channels formed in
reconstitution experiments had a conductance of 7.6 nS in 1 M KCl. Both
Lpg1974 proteins formed pores that were voltage-dependent and
anion-selective similar to the pores formed by hVDAC1. These results
suggest that Lpg1974 of L. pneumophila is indeed a structural and
functional homologue to hVDAC1.
Esna Ashari Z, Dasgupta N, Brayton KA, Broschat SL.
PLoS One. 2018 May 9;13(5):e0197041. doi: 10.1371/journal.pone.0197041. eCollection 2018.
A thorough literature search was performed to find features that have
been proposed. Feature values were calculated for datasets of known
effectors and non-effectors for T4SS-containing pathogens for four
genera with a sufficient number of known effectors, Legionella
pneumophila, Coxiella burnetii, Brucella spp, and Bartonella spp. The
features were ranked, and less important features were filtered out.
Saleem F, Mustafa A, Kori JA, Hussain MS, Kamran Azim M.
Microb Ecol. 2018 Apr 24. doi: 10.1007/s00248-018-1192-2. [Epub ahead of print]The present metagenomic analysis of DWSS of Karachi has allowed the
evaluation of bacterial communities in source water and the water being
supplied to the city. Moreover, measurement of heavy metals in water
samples from Karachi revealed arsenic concentration according to WHO
standards which is in contrast of recent study which reported extensive
arsenic contamination in aquifers in the Indus valley plain. To the best
of our knowledge, this is the first metagenomic study of DWSS of
Karachi.
Ghilamicael AM, Boga HI, Anami SE, Mehari T, Budambula NLM.
PLoS One. 2018 Mar 22;13(3):e0194554. doi: 10.1371/journal.pone.0194554. eCollection 2018.
Human pathogens can survive and grow in hot springs. For water quality
assessment, Escherichia coli or Enterococci are the main thermotolerant
enteric bacteria commonly used to estimate the load of pathogenic
bacteria in water. However, most of the environmental bacteria are
unculturable thus culture methods may cause bias in detection of most
pathogens. Illumina sequencing can provide a more comprehensive and
accurate insight into environmental bacterial pathogens, which can be
used to develop better risk assessment methods and promote public health
awareness. In this study, high-throughput Illumina sequencing was used
to identify bacterial pathogens from five hot springs; Maiwooi, Akwar,
Garbanabra, Elegedi and Gelti, in Eritrea. Water samples were collected
from the five hot springs. Total community DNA was extracted from
samples using the phenol-chloroform method. The 16S rRNA gene variable
region (V4-V7) of the extracted DNA was amplified and library
construction done according to Illumina sequencing protocol. The
sequence reads (length >200 bp) from Illumina sequencing libraries
ranged from 22,091 sequences in the wet sediment sample from Garbanabra
to 155,789 sequences in the mat sample from Elegedi. Taxonomy was
assigned to each OTU using BLASTn against a curated database derived
from GreenGenes, RDPII, SILVA SSU Reference 119 and NCBI. The proportion
of potential pathogens from the water samples was highest in Maiwooi
(17.8%), followed by Gelti (16.7%), Akwar (13.6%) and Garbanabra
(10.9%). Although the numbers of DNA sequence reads from Illumina
sequencing were very high for the Elegedi (104,328), corresponding
proportion of potential pathogens very low (3.6%). Most of the potential
pathogenic bacterial sequences identified were from Proteobacteria and
Firmicutes. Legionella
and Clostridium were the most common detected genera with different
species. Most of the potential pathogens were detected from the water
samples. However, sequences belonging to Clostridium were observed more
abundantly from the mat samples. This study employed high-throughput
sequencing technologies to determine the presence of pathogenic bacteria
in the five hot springs in Eritrea.
Kubori T, Kitao T, Ando H, Nagai H.Cell Microbiol. 2018 Jul;20(7):e12840. doi: 10.1111/cmi.12840. Epub 2018 Apr 6.... Many effector proteins are expected to be involved in biogenesis and regulation of the Legionella-containing vacuole (LCV) that is highly decorated with ubiquitin. Here, we identified a Legionella deubiquitinase, designated LotA, by carrying out a genome
analysis to find proteins resembling the eukaryotic ovarian tumour
superfamily of cysteine proteases. LotA exhibits a dual ability to
cleave ubiquitin chains that is dependent on 2 distinctive catalytic
cysteine residues in the eukaryotic ovarian tumour domains. One cysteine
dominantly contributes to the removal ofubiquitin from the LCVs by its
polyubiquitin cleavage activity. The other specifically cleaves
conjugated Lys6-linked ubiquitin. After delivered by the Type 4
secretion system, LotA localises on the LCVs via its PI(3)P-binding
domain. The lipid-binding ability of LotA is crucial for ubiquitin
removal from the vacuoles. We further analysed the functional
interaction of the protein with the recently reported noncanonical
ubiquitin ligases of L. pneumophila, revealing that the effector
proteins are involved in coordinated regulation that contributes to
bacterial growth in the host cells.
Genome Announc. 2018 Feb 1;6(5). pii: e01588-17. doi: 10.1128/genomeA.01588-17.Legionella sainthelensi
is an aquatic environmental bacterium that in humans can cause
Legionnaires' disease (LD), an often severe form of pneumonia. Here, we
report the first complete genome of a L. sainthelensi clinical isolate obtained in 2001 from a patient with LD in Canterbury, New Zealand.
Currently, there are two draft genome sequences for the ATCC 35248 type
strain, which are the environmental isolates obtained near the Mt. St.
Helens volcano (GenBank accession numbers NZ_JHXP00000000 and NZ_LNYV00000000).
Highlights Previously, it was demonstrated that L. pneumophila consists of three subspecies: pneumophila, fraseri and pascullei.
A set of 38 complete L. pneumophila genomes was analyzed. Four distinct subspecies, including the novel subsp. raphaeli, were identified. ANI values show unusually large genomic distances among subspecies. Subspecies-specific SBT consensus patterns were determined. A panel of subspecies-unique genes can be used for testing and classification.

 ...
...